As the physical anthropologist seeks to understand the meaning of the striking range of variability which confronts him he is constantly faced with the importance of the processes of physical growth and development. In addition to the obvious fact that the size and shape of the adult are the direct result of his own unique pattern of development, there are a number of other ways in which ontogeny (i.e., development) is responsive to the evolutionary forces which affect human populations. For example, the longer growth period of Homo sapiens relative to other Primates (successively longer periods being a feature observed as we proceed up the Primate scale) is of selective value in affording more time in which to learn the many facets of human culture while still socially and biologically dependent upon one’s parents. The study of growth and of the forces which which create and maintain its variability are basic parts of any discipline which seeks to understand man in his entirety. Small wonder it is, then, that anthropologists such as Boas, Krogman and Garn have contributed many of the basic advances in growth research, while other biological scientists, such as Todd, Skerlj and Tanner, who have likewise contributed much, have always been closely associated with anthropology. In other words, though human growth research has always been multi-disciplinary in nature, anthropologists have been prominent throughout its rather short history.
One of the most striking features of growth is its regularity. If we plot repeated measurements of, for example, the height of a single child as taken by a single trained measurer against the age of that child, as shown in Figure 1, we will immediately note the smoothness of the resulting curve. In fact, it is so smooth that it may be accurately described by a very simple mathematical equation. Even the course of adolescence, characterized by a rapid and intense spurt in growth, proceeds along a smooth pathway for the same individual. The regularity in the shape of the growth curve is such that, as early as five years of age, we may estimate the adult height of a particular child and be confident of accuracy within a range of about two inches on either side of our estimate.
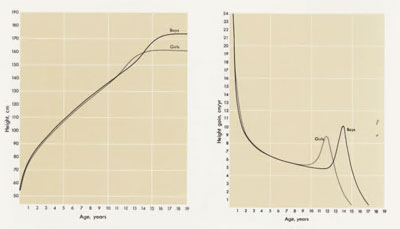
Growth may be characterized as “target-seeking,” a description which is the more apt due to the trajectory-like shapes of the curves of most dimensions. The target is, of course, the adult and it is sought unerringly and constantly. The pathway to this target is apparently built into the genetic code and, when it is disrupted, corrections are made to allow the resumption of the original trajectory. Thus, when a healthy child suffers from a particularly acute illness, or is beset by some prolonged disease or chronic upset, his growth progress is usually affected and he falls off course, usually, but not always, dropping behind. When the disease is cured, or the condition corrected, growth resumes at a faster-than-usual pace and the individual is observed to “catch up.” The period of acceleration continues until the child reaches the approximate position on the curve that he would have been at had the disruption not occurred; at that point, the growth slows down to its normal rate.
Just as we may characterize growth as regular and predictable, we may also characterize it as highly variable. Such an apparent paradox may easily be resolved. The regularity and predictability refer to any single dimension, such as height, in a single child, while the variability refers to the relationship between two dimensions in the same individual, such as height and weight, or between the same dimension in two or more individuals. For example, during the adolescent phase of growth in males, there is a loss of subcutaneous fat in both the arm and the leg. Before adolescence, a boy who has a thick layer of fat on his arm will quite likely have a thick layer in his leg. However, there is very little relationship between the timing of this loss in the two extremities of an individual boy. In one person, the loss may occur first in the leg, then later in the arm; in another, this may be reversed; in yet a third the loss may occur simultaneously in both members. In other words, all possible combinations of variability can be seen in normal, healthy boys and the precise sequence cannot be predicted in a specific one with any confidence at all. The range of variability among individuals with respect to a single dimension is likewise striking and, in a sense, distressing. Twenty-five years ago, researchers were very much concerned with the construction of “standards” of growth, which would provide the reference norms for sizes of children, which could be applied to a wide variety of children and which could pinpoint the abnormal and, hence, unhealthy growers. We now know that such standards are of little value except in restricted groups and, often even then, serve more as a series of flexible guidelines. We can easily demonstrate that the growth of normal white children from Des Moines or Cleveland is not representative of the growth of normal white children from Philadelphia. If we desire to learn how a particular child is growing relative to a meaningful norm, then we must use, as our standard, the growth pattern of his or her peers, from the same group and representing the same unique combination of ethnic elements and environmental factors characteristic of any single population segment. Even when we do have adequate and representative standards of normal growth, the range of variation is so wide that we must be very conservative in our evaluations of the individual or group. Such a range of normal variation is a remarkable feature of our species and is indicative of both the many forces which can affect growth and the extreme plasticity of the growth process in the face of these forces.
The growth status of a child (or group of children) at any given moment in time is controlled intrinsically by two sets of factors: those which relate to his final adult size and those which relate to the speed at which that size is being attained. Thus, it is almost without exception that a tall adult was a tall child, and in about the same relative position. It is this feature which allows us to predict the adult height of a child by the age of five with a reasonable degree of accuracy for some purposes. If a five year old boy is in the tallest ten percent for his age, the chances are he will be in the tallest ten percent as a man (though his relative position may change somewhat during adolescence). The regularity of the growth pattern of individuals results in the “canalization” of development and adult size emerges as a very powerful determinant of size throughout growth.
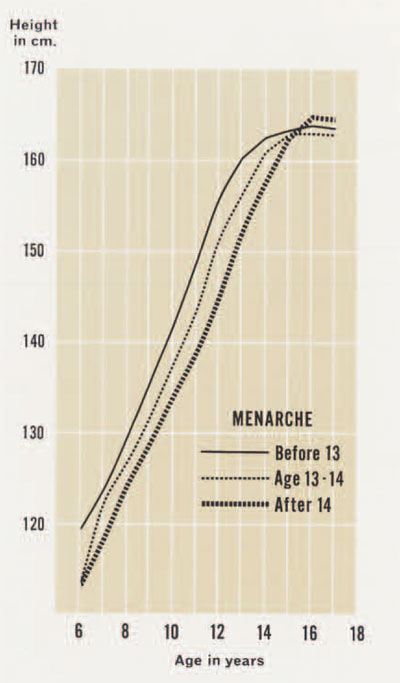
On the other hand, children vary considerably in the rate at which they move toward the adult. The average American boy will grow for eighteen years; however, some will take twenty years and others but sixteen to reach adult size. Called the rate of maturation, this feature is under direct genetic control, though subject to environmental forces, and the variability will affect the size at a particular age. Other things being equal, the faster maturers of a given age (say ten years) will be larger than the slower maturers. This is because a faster maturing ten year old is closer to the adult than a slower maturer. Variability in the rate of maturation is most notable during adolescence. A rapid maturing ten year old girl will be well into her adolescent spurt, perhaps even at her peak velocity, and will therefore be significantly larger than a slow maturing one, who will still be growing at the much-reduced preadolescent speeds. The degree of this difference is illustrated in Figure 2, which shows the curves of growth in height for samples of fast, average, and slow maturers. In this case, assignment to a maturation rate group is made on the basis of the age at menarche, a very sensitive indicator of such differences. It is clear that the same relationships exist among the three groups throughout the period of growth, though the differences become intensified during adolescence. It is interesting and important to note that these differenced disappear when adulthood is reached; the maturation rate does not really affect the final height of an individual in a significant way but it does add to the variation observed during growth.
The interplay of these two determinants, size and maturation rate, is very complex but must be analyzed if we are to draw meaningful conclusions from our data. Thus the comparison of children of the same chronological age has very little, if any, meaning since we can easily be misled by size differences which are due to differences in their rates of maturation. Simply stating that a particular group of twelve year old boys is larger than another group of the same age tells us nothing unless we have information on their respective rates of maturation. If, on the other hand, we can state that the one group is maturing more rapidly than the other, we have established a more basic difference. Then, even after controlling for this maturation rate differential, if we are able to conclude that, in our samples, those boys of the one group are larger than those of the other of equivalent levels of maturation, we have begun to make a meaningful analysis of the differences between our groups.
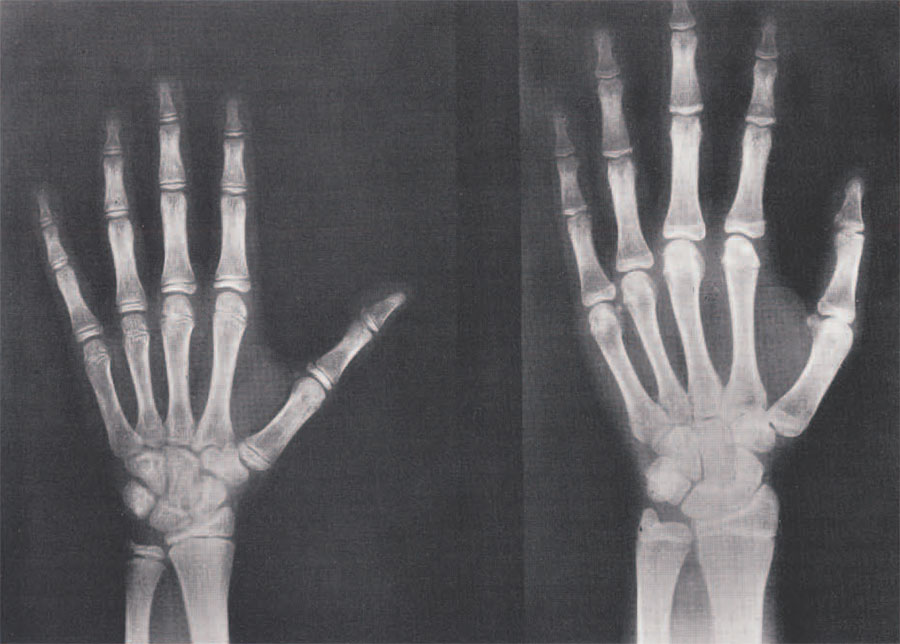
The most suitable technique presently available for allowing us to determine the rate of maturation in children is based upon changes in their skeletons, which we can view on appropriate X-rays. These changes involve, 1) the initial appearance on the film of particular bones, signifying the beginning of their conversion from cartilage to bone; 2) the shaping of bones to their adult forms through the addition of bony tissue at some sites and the removal at others; and 3) the fusion, in the long bones, of their ends, or epiphyses, with their respective shafts. The rate at which these changes take place accurately reflects the overall rate of biological maturation and, when skeletal maturation is complete, we find that growth in size has likewise ceased. Figure 3 shows the x-ray views of the hand skeletons of two girls, illustrating variation in skeletal maturation at the same age, with concomitant variation in biological maturation as well.
The other important intrinsic determinant of growth status is sex, since basic differences in the shape and timing of the growth curve exist between males and females. More rapid maturation of the female is characteristic of a number of mammalian species. In man, it is on the order of two years; girls commence their adolescent spurt about two years prior to boys, they reach puberty two years sooner, on the average, and they complete growth some two years before males. Before adolescence, this makes very little difference, since boys and girls do not differ significantly in size during these ages. However, since girls enter into their growth spurt some two years earlier, there is a period in which they are taller than boys. However, boys eventually exceed them in height and retain this differential as adults.
It is significant that the actual magnitude of the adolescent spurt has but a minor role in the genesis of size differences between male and female adults. The major factor in the greater heights of males is the additional two years of growth just prior to the onset of their adolescent spurt, when the greater height of the male is established. The magnitude of the adolescent spurt is about the same in both sexes, boys putting on only a bit more height during this period.
Sex differences also exist in the growth of the various “components” of body mass, notably fat, muscle, and bone. Prior to adolescence there are but small quantitative differences in the amounts of these tissues, though boys tend to be somewhat more muscular and girls have small, but consistently greater amounts of fat. However at adolescence, presumably due to differential patterns of hormone secretion, profound differences emerge. Girls continue to accumulate fat on both their trunks and their extremities at rates approximately equal to those of pre-adolescence. Boys display such a continuation as far as fat on their trunks, but undergo a marked loss of extremity fat, about one-half of the maximum value before adolescence. At the same time they undergo a marked spurt in muscle mass. The results, which persist into adulthood, are that some twice the percentage of body weight is fat in females, compared to males.
Unfortunately, our “anthropological” knowledge of growth is relatively sparse and we cannot make the broad cross-populational comparisons that we can for other biological systems. The majority effort, even among anthropologists, has been devoted to determining the patterns of growth in European or North American whites, analyzing the variability and studying the effects of inherited and acquired aspects which affect size and rate of maturation. In addition to this simple lack of available data, we are also confounded by the profound alterations of growth patterns due to unfavorable environmental conditions. The startling deficiencies in the nutritive intakes of many populations, as well as differences in their genetic structures, render it difficult to make any really meaningful comparisons. How can we discuss differences in the basic growth patterns between American whites and Papuans when the former are both under-exercised and over-fed and the latter exist often on less than 2,000 calories per day? Or how can we speak of an inherited pattern of development among Gambian children who suffer chronic and acute episodes of parasite infestation during the rainy season? Assuming that real genetic differences in growth may exist, how can we even be certain that the differences that we observe under such conditions are, in fact, due to environmental effects? We are still involved in trying to separate those components of variation which are du to the environment from those due to the hereditary background of the population.
Despite the above difficulties, we can make some statements about population differences in growth and evolutionary trends in development. For instance, we now know that American Negroes are, at birth, both larger and biologically more mature than whites. This differential persists through early childhood, but disappears at about six years, with some evidence that Negroes enter adolescence at a later age than do whites. We are certain that this advancement has a genetic basis; we are not sure about the later slowdown, since it has been shown that the diets of many American Negroes are less adequate than those of whites. It is quite possible that this lag is due to the cumulative effects of a less optimal environment.
We also know that for all populations on which we have data children of one generation are maturing more rapidly than are those of the preceding one and are becoming larger as adults. The increase in the rate of biological maturation is such that girls of each new generation are reaching puberty about three months earlier than their mothers. In parts of Scandinavia, where long-term records are available, these trends are clear and unmistakable from the mid-nineteenth century to the present day. The same is true, though the time depth is not so impressive, for the United States, Great Britain, Poland, and Czechoslovakia. The cause of this “secular trent” is not completely determined as yet, with proponents of improved environment on one side and those of evolutionary changes on the other. In favor of the former is the fact that adverse environments hit the maturation rate very hard while improved health and nutrition have always resulted in more rapid growth. In favor of the latter are the data which show that the secular trend is occurring in the upper income groups at about the same intensity as in the lower, who would presumable have the most to gain from environmental improvement. J.M. Tanner, who has studied this phenomenon closely, feels that the increase in the rate of maturation is probably the increase in the rate of maturation is probably the result of improved nutrition, medical care, etc. However, the trend toward greater height in adults may very well be an evolutionary change.
I have, in this short paper, tried to show the directions in which the anthropological study of growth is proceeding. I have omitted, largely for lack of space, considerations of physiological and endocrinological aspects of growth, as well as any mention of changes in shape or proportion. I fear that I have raised as many problems as I have solved but perhaps this is the nature of science. Certainly it is the case in studies of human development which are, one must remember, still relatively new and still in the process of methodological refinement and anthropological application.