Who are the Polynesians? From whom did they derive? How were they able to reach the far-flung islands of the extreme eastern Oceanic region? These are the questions that have both vexed and intrigued scholars and students of Pacific culture since the discovery of the Pacific islands.
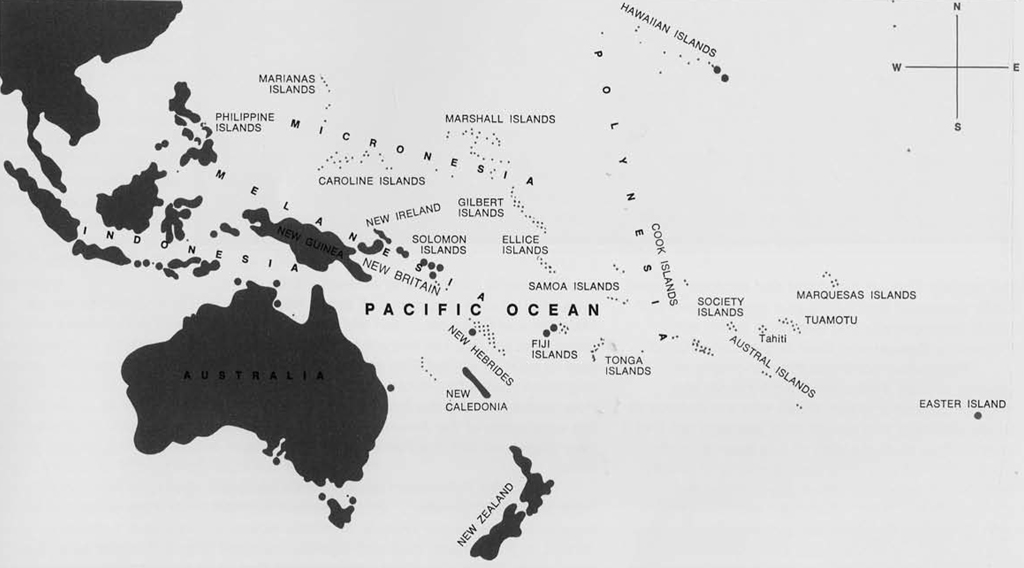
As intrepid European explorers began venturing into the newly discovered Pacific during the latter half of the 17th and early 18th centuries they found the countless islands and archipelagos inhabited by seemingly diverse groups of people. By the middle of the 19th century the islands of Oceania had been divided into the discrete geographic areas of Polynesia, Melanesia, and Micronesia—the boundaries of which were based primarily on the outward manifestations of race, language, and the customs of their indigenous populations.
Of the three geographic regions mentioned above, the Polynesian islands have held an enduring fascination for western man since their discovery, and it was perhaps inevitable that the tall, brown-skinned Polynesians would receive the brunt of European missionization, exploitation, and academic study. From a 19th century European point of view the Polynesians were physically attractive, generous, and receptive—in contrast to their black, short-statured Melanesian neighbors to the west. As contact with Polynesia by early explorers and missionaries increased, conjecture concerning the origin and migration of this widely dispersed “race” began to follow a number of paths.

Mistakes have been made in past studies by assuming that Polynesian race, culture, and language—as wholes—were homogeneous to the extent that information gathered in one island could be applied equally well to all other islands within the Polynesian geographic area commonly known as the “Great Triangle.” As early as 1784 the famous English navigator and explorer James Cook proposed the outline of a general theory of racial homogeneity and language purity within Polynesia. Concerning their origin Cook stated:
“From what continent they originally emigrated, and by what steps they have spread through so vast a space, those who are curious in disquisitions of this nature, may perhaps not find it very difficult to conjecture. It has been already observed, that they bear strong marks of affinity to some of the Indian tribes, that inhabit the Lad rones and Caroline Islands; and the same affinity may again be traced amongst the Battas and the Malays. When these events happened, is not so easy to ascertain; it was probably not very lately, as they are extremely populous, and have no tradition of their own origin, but what is perfectly fabulous; whilst, on the other hand, the unadulterated state of their general language, and the simplicity which still prevails in their customs and manners seem to indicate, that it could not have been at any very distant period.”

In a rather tortuous way, Cook reasoned that the large population and the lack of any tradition explaining their origin indicated a long period of island occupation. On the other hand, the purity of their language and simple customs indicated a comparatively short period of occupation. Hence the inherent contradictions of the problems were established and recognized.
Early justifications for the racial identity of the Polynesians—as distinct from Melanesians—ranged from Samuel Marsden’s postulation in 1819 that the Polynesians were intimately linked with the lost tribes of Israel as recorded in the Old Testament, to William Ellis’ observations in 1830 that the Polynesians held many similarities with the people of the Malay tribes. Basing his case primarily on linguistic evidence, Ellis adhered to the view of a common origin not only for Malayan and Polynesian, but the American Indian languages as well. His knowledge of prevailing east to west winds led Ellis to hypothesize that migration routes had taken the ancestors of the Polynesians across the Bering Straits down along the west coast of the American continent, where they dispersed into the Pacific basin. Quoting Ellis:

“If . .. [the Polynesian islands] were peopled from the Malay Islands … [the Polynesians] must have possessed better vessels than they now exhibit, to have made their way against the constant trade-winds prevailing within the tropics, and ,blowing regularly, with but transient and uncertain interruptions, from east to west….”
As an alternative to Ellis’ American origin of east-west migration and Cook’s Asiatic origin of a west-to-east migration, Moerenhout concluded in 1837 that the Polynesians should be considered autochthonous to Oceania because the uniformity of their customs and language could only be accounted for by the existence of a once-great Oceanic continent that had subsided, leaving only the highest mountain peaks thrusting their heads above the waters of the Pacific. Although Moerenhout’s reasoning was later disproved as geological knowledge increased, the “sunken continent” theory received serious consideration and was not unsophisticated in its time.
Thus, as evidence accumulated rather slowly (evidence that was at best indecisive) a continuing elaboration of the three theories outlined above was carried out until John Fraser’s postulation in 1895 of a series of “waves” of immigrants into the Oceanic basin. Fraser felt that distinct racial elements, migrating at different times, had settled the Oceanic islands, and in the process had become mixed. Fraser’s theory was rather complex:

“My explanation of the whole matter under discussion is briefly this: the main officina gentium for Oceania, long, long ago was India. The whole extent of that peninsula was at a very early period probably more than twenty centuries before the Christian era, occupied by a pure black race, which I call Hamite; later on, there came into it a Cush ite race, also black, but more mixed than the Hamites…. In Eastern Polynesia, the aboriginal black population must have been very scant, as these islands are so far removed from the Asian continent, and consequently the traces of their occupation have been swamped by the subsequent flow of Polynesian immigrants…. Fiji also has two black races, those of the interior and those of the coast, and these show important differences in customs; so also in New Guinea…. In fine, I think it could be established with the utmost probability that two black races, proceeding from India in succession, peopled the whole of the islands of Oceania.”
Fraser was positive that in certain isolated areas, pure racial stocks could be recognized which would substantiate his theory of mass migration.
As our knowledge of Polynesia increased, it became ever more difficult to justify a theory of homogeneous physical type for the Polynesian people. Until the turn of the century little work had been conducted under the guise of physical anthropology. In 1907, J. Macmillan Brown was the first scholar to attempt an interpretation of Polynesian origins and migrations based on physical observations. In 1914, Friedrici, utilizing data from both physical anthropology and linguistics, suggested that there were three basic racial elements to be considered in theories about the settlement of the Pacific basin. Not until the late 1920’s was a concerted attempt made to record the metrical data of living populations within Polynesia and Oceania.

The work of Roland B. Dixon and Louis R. Sullivan began to lay the foundations for a highly sophisticated study of physical anthropology in the Pacific. Sullivan, addressing the First Pan-Pacific Scientific Conference in 1920, remarked that “the data is entirely inadequate for conclusions as to the racial or inter-insular affinities of the Polynesians.” Sullivan felt that physical anthropology “could not offer a solution to the problems of origin and migration” but could describe the Polynesian groups accurately. He felt that metrical data could prove beyond a reasonable doubt the racial affinities of the Polynesians and designate the racial branch to which they belonged. In order to equate the great physical diversity within Polynesia, Sullivan proposed four waves of racially distinct elements. He considered two of the races to be Caucasoid, one Negroid (Melanesian), and a fourth, with doubtful affiliation, to be a combination of Negroid and Mongoloid characteristics. The four separate types, he felt, had mixed in varying proportions to make up the composite populations of the various island groups. Sullivan touched lightly on the possibility of physical differentiation through the process of isolation within Polynesia, but came to the conclusion that “the distribution of types both within and without Polynesia argues strongly against a local origin of these types in Polynesia.”
In a short article entitled “A New Theory of Polynesian Origins,” Roland B. Dixon analyzed cranial measurements from living populations in Oceania and Southeast Asia and concluded that four racial types were represented in Polynesia. Dixon used as his criteria cephalic, length-height, and nasal indices. He made the assumption that those groups whose indices were extreme on either end of the several series constituted fundamental types, while those falling closer to the median values were racial blends. He suggested in a later view that five waves of racially distinct people were required to people the far reaches of eastern Polynesia and specifically Easter Island.
In a recent assessment of data relevant to Polynesian origins, Kenneth P. Emory of the Bernice P. Bishop Museum in Honolulu has proposed the following view:

“It seems … erroneous to consider that there was a migration to Polynesia of a people physically identical with the Polynesians as we know them, and as already possessing the distinctive features of Polynesian language and culture. What now appears most likely is that people of somewhat diverse origins came together in a western archipelago in the Polynesian area about 1500 B.C., and in comparative isolation, their descendants, their language, and their culture took on the features which Polynesians now share in common and which give them their distinctive characteristics.”
Emory’s view of Polynesian origin is currently shared by many Pacific scholars and has been elaborated by Roger Green. Based on present linguistic and archaeological evidence, Green feels that the western archipelago proposed by Emory is to be identified as Fiji, and that the evidence for a Melanesian origin of the Polynesians in Fiji is fairly straightforward.
If the immediate origins of the Polynesians are to be sought in Melanesia, as Emory, Green, and others have suggested, we must then ask what factors have caused the apparent racial diversity and assumed cultural disparity between the two groups, i.e., Melanesian and Polynesian?
During the past twenty or so years there has been a general concensus among scholars that there is some relationship between the geographical environment and the physical manifestations of its inhabitants. This view has been cogently argued by Carleton Coon, Marshall and Snow, Harrison and his associates, and others. Pacific studies of the past have generally disregarded the ecological factors present in the Pacific because they were thought to be uniform, or generally so, throughout the entire area. From recent geological and ecological studies the Pacific islands have been found to be quite diverse.
The Pacific islands are spread out over more than sixty million square miles of ocean, most of them lying within the tropical zone. This vast area is composed of at least four basic island types:
Continental islands such as New Zealand and New Guinea.
- The high volcanic islands of Hawaii, Fiji, and the Society group.
- Low, non-volcanic islands composed of tuff superficially covered with coral and foraminiferous limestones.
- Atolls, consisting of coral reefs inclosing lagoons and supporting low, sandy islands.
Even while we recognize their great geographic separation and environmental diversity, each island group within Oceania may be loosely considered an almost discrete or closed micro-environment. Ecological variation will effect differences in the dietary options available to some island inhabitants in contrast to the options available to other island groups. According to A. P. Vayda, material prosperity, the ability to continue established tradition or actualize new possibilities. social organization, stratification, population density, and breeding habits are all directly or indirectly affected by the environment.
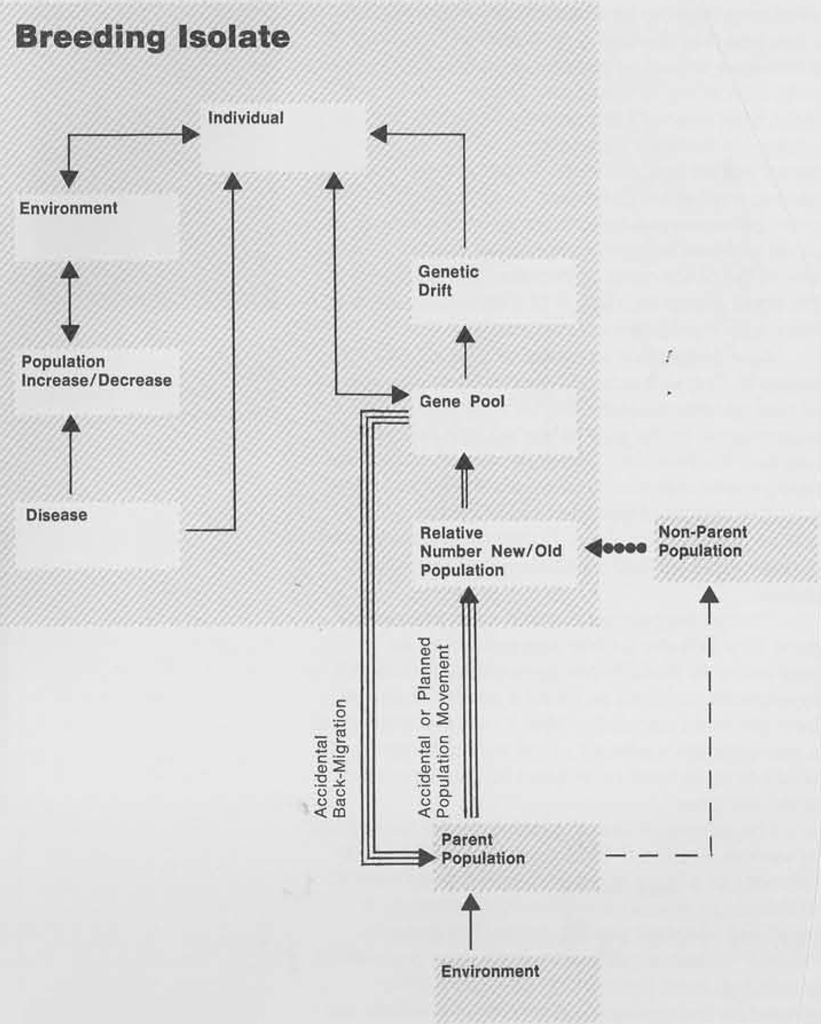
Of equal importance in affecting population differentiation is the number of original colonizers on an island and the particular segment of a larger parent gene pool that they represent—that segment being what is operating in the particular island environment. The subject of colonization and voyaging in the Pacific islands has been controversial since their discovery. In a review in 1956 of the achievements and deficiencies of pre-European Polynesian voyaging, Andrew Sharp argues that Polynesia was peopled as a result of “accidental” landfalls of voyagers lost at sea or sent into involuntary exile. Before Sharp’s study the orthodox view held by most anthropologists in Oceania was that pre-European Polynesians were capable of maintaining regular contacts between islands separated by 2,000 miles of open ocean, and that the infiltration and subsequent settlement of Polynesia resulted from voyages of “exploration and large-scale migrations” to the newly discovered lands. Sharp’s study of the ethnographic resources available for Polynesia did not support this view. It suggested a theory of initial “accidental” discovery and settlement from west to east, followed at a later time by further “accidental” discovery and settlement along the prevailing lines of drift from east to west. Concerning the effect of such population movements on the genetic and physical characteristics of small populations, Ward Goodenough states:

“Given the accidental settlement of Polynesia and Micronesia by a few small groups of occasional west-to-east castaways and a much larger number of their descendants as later east-to-west castaways, and given the progressive genetic isolation from west to east by way of initially very small populations which this implies, we should expect to find increasing homozygosity from group to group the further east we go into Oceania from Melanesia.”
Goodenough has further pointed out that tradition indicates a great deal of back-and-forth movement within the western Polynesian area of Fiji, Tonga, Samoa, etc., while the implication for eastern Polynesia would indicate that once settled, the islands remained as a whole separated from the rest of the Pacific.
Recent blood group and genetic studies of Melanesian populations have led Swindler and Simmons to conclude that the Melanesian populations are sufficiently homogeneous to be regarded as a major breeding unit or geographic race. The constant exchange of genetic materials within a geographically delimited area over a significant period of time has made the various groups within Melanesia more like each other than they are like groups from outside the area. These findings would indicate then, that a great deal of change has taken place within the area, with the likelihood that local populations have undergone major genetic and morphological changes as compared with the initial settlers from whom they may be derived. B. Glass has stated that if a segment of the gene pool were isolated from the larger population, the manner and degree in which the frequencies within the “new” isolate gene pool would depart from the prevailing larger unit would exclude certain potentialities and, by this exclusion, increase the probability that other potentialities would be actualized. It has been found by Wright that in very small isolated communities, minor fluctuations in gene frequencies rarely if ever contribute to an evolutionary advance but often result in marked differentiation by random processes known as genetic drift. Random genetic drift, according to Glass and Brues, can do little more than intensify the genetic variability that is present in isolated populations, while the same factor will be ineffective on larger populations because selective pressures continue to act with great constancy. A later increase in the size of a small isolated breeding population will not reverse any of the tendencies or peculiarities acquired by the small group as a result of drift but will convert them into stable characteristics.

As a population expands in number and continues to drift in a closed breeding unit, the impact of new genetic material on the population will be proportional to the size of the established population and the relative number of new arrivals. If the new genetic material arises from the parent gene pool the effects of genetic impact on the isolate will be greatly minimized. Concerning the input of genetic material into a population isolate Glass states:
“It may be pointed out that even though the gene flow into the isolate was estimated as amounting to 10-15% per generation, that is still in absolute numbers so small a supply of genes from the main population that it could scarcely be a representative sample of the surrounding gene pool but must itself be subject to the vagaries of small samples.”
The effects of Glass’ argument may be applied in several ways to the Pacific islands. The establishment of a fairly large population in a volcanic archipelago would be minimally affected by a small but constant genetic input. The genetic “stamp” of the larger, well-established population would not, in all probability, be significantly altered as it absorbed new arrivals. However, on very small islands or coral atolls one canoe-load of new arrivals could drastically alter the genetic resources within one generation. Since land and food resources are considerably restricted in low islands, maximum population expansion would be considerably smaller than in the thickly soiled volcanic areas. An expanded atoll population could be substantially reduced by the exigencies of tidal wave, famine, or typhoon within a very short period. The effect of such an occurrence has been documented by the devastation of Mokil atoll in 1775 by a typhoon, leaving only 25 or 30 survivors. These considerations then would imply that the numerical disparity between local populations and parties of new arrivals (whether accidental or not) was generally likely to be much smaller in coral atolls than in volcanic islands. A. P. Vayda has cogently argued this point, using the island of Niue as an example:

“Niue is visited by hurricanes about once in ten years, and a drought occurs nearly every year. . .. The people … were always subject to periodic famine and devastation…. Accordingly there may have been several major population reductions in the course of Niuean prehistory. Two suggestions, first that Niuean population may sometimes have been reduced to much smaller size … [and] second, that this smaller size may have been such that any relatively large parties of accidental voyagers would have made contributions to Niuean culture and genetic makeup discernible in the ethnographic and phenotypic present.”
The significance of external disease factors affecting Oceanic population numbers is of utmost importance in relation to the measurement and documentation of living peoples. Of the population found during early contact times it has been estimated that only approximately one-tenth survived the cataclysmic effects of smallpox, venereal disease, tuberculosis, measles, and the common cold introduced by traders, sailors, missionaries, and colonial government officials. Coupled with a marked decline in population was the influx of new genetic resources from the disease carriers mentioned above. Most of the highly developed aboriginal centers were hardest hit by disease and most affected by European genes because of their desirability to Europeans as trade centers and as areas with good land resources. Mass depopulation of major population centers left many “back-water” areas that were relatively free of traders but that were later primary targets for the missionary societies of Europe, America, and Australia. European genes during post-contact times were, in many cases, unequally distributed by social class. While Asian and Indian genes were more-or-less restricted to the less socially prominent families, Europeans mated freely with persons of nigh rank. Plantation laborers brought back alien wives of Oriental and Australian aboriginal background to further alter the genetic make-up of island populations.
Physical types could hardly be expected to remain unaltered, especially in view of the severe decline in individual island populations that has taken place. From these congeries new breeding populations were formed, and although a more-orless common cultural base was carried from island to island, these social forms underwent change in much the same way as genetic characteristics—becoming adjusted to new and changed environmental conditions—adapting through a process similar to genetic micro-evolution. Each island population has developed historically its own distinct cultural and genetic direction to produce the Polynesian people as a distinct entity amongst the races of Man. As the geneticist Simmons has stated, “… there is no Polynesian problem other than that created by ourselves.”